








Por: Ángel Humberto Rojo-Cerebros y Mirian Celene León-López*
El cultivo de la lobina se encuentra poco desarrollado en México, a pesar de que existe alta demanda para consumo humano y de ser el pez más importante de pesca deportiva continental; quizá debido al mito de que en cautiverio presenta un factor de conversión alimenticia (FCA) exageradamente alto. Sin embargo, la presente evaluación indica que la lobina tiene la capacidad de aprovechar eficientemente los alimentos administrados y el supuesto alto FCA es solo un mito.
Introducción
La lobina de boca grande (largemouth bass, nombre común en inglés) Micropterus salmoides (Lacépède, 1802) pertenece a la familia Centrarchidae y es el pez dulceacuícola más importante de pesca deportiva. Es nativo de América del Norte y ampliamente introducido en Europa, África, América del Sur, Japón y China.
Cabe señalar que también es un pez de importancia acuícola, en el 2017 la producción mundial piscícola de lobina fue de 458,000 toneladas. La mayor parte de la producción fue de China (99.8 %) donde es muy valorado como alimento (Hussein et al., 2020).
“En los Estados Unidos de América (EUA), 31 estados cultivan y producen lobina, con ventas que alcanzaron los 27 millones de dólares americanos en 2018.”
Además, en América del Norte, la lobina boca grande es un pez muy valorado como alimento para la comunidad asiática, en especial en las grandes ciudades como Nueva York, Chicago y Toronto, lo que despierta mucho interés en su producción acuícola.
Las lobinas pertenecen al género Micropterus que consta de ocho especies, pero la comunidad pesquera y acuícola está más familiarizada con dos, la lobina boca grande (Micropterus salmoides) y la lobina de boca chica (M. dolomieui).
Sin embargo, hay dos subespecies reconocidas de lobina boca grande: la lobina boca grande norteña, M. salmoides salmoides y la lobina boca grande del sur o Florida, M. salmoides floridanus (Glenewinkel et al., 2011). Las subespecies y sus híbridos solo se pueden distinguir mediante pruebas genéticas (Villa-Melchor, 2016).

El factor más importante en la especiación de M. salmoides es la adaptación a las temperaturas en las latitudes que se distribuye, por ejemplo, en la latitud del sur de Illinois, la subespecie Florida suele morir en el invierno en estanques poco profundos, lo cual no ocurre con la subespecie norteña (Heidinger, 2000).
En México, a los individuos de las subespecies de M. salmoides se les llama comúnmente lobina negra, y son de mucha importancia como recurso pesquero para las comunidades de pescadores y para el mantenimiento de la pesca deportiva-recreativa (Beltrán-Álvarez et al., 2013).
Aunque cabe señalar que los estudios de genética poblacional de lobina negra en los cuerpos de agua continentales en México han reportado un marcado dominio en presencia de la subespecie de Florida sobre la subespecie norteña, y que además presentan ejemplares con mejores tallas y pesos (Villa Melchor, 2016); la anterior sin duda como resultado de la preferencia de la subespecie Florida a temperaturas más cálidas que prevalecen en los cuerpos de aguas continentales de México.
Las experiencias piscícolas con lobina que aquí se describen se llevaron a cabo en las instalaciones de Dunn´s Fish Farm Inc. que se encuentra en el estado de Arkansas, EUA; compañía que tiene como giro comercial la producción a ciclo cerrado de la lobina boca grande norteña (largemouth bass) en presentación entera para el consumo humano.
Aunque en diversas ocasiones los juveniles se destinan a la repoblación de lagos y ríos, con fines de pesca deportiva y, en otros casos, para la investigación acuícola.
Retos en la producción de juveniles de lobina
Aunque se han desarrollado técnicas de reproducción y cultivo larvario de lobina en criaderos, existen desafíos relacionados con impredecible fecundidad, mala calidad de huevos, mortalidad masiva de larvas y deformidades que se presentan desde el desarrollo ontogenético temprano.
Una alternativa para mejorar la producción y calidad de juveniles de peces incluye el enfoque dietético de los reproductores porque muchos nutrientes necesarios para un desarrollo temprano adecuado se transfieren en los huevos por vía materna (Izquierdo et al., 2015).
“Esto nos motivó a realizar un estudio piloto para evaluar el efecto de la suplementación dietética de aceite de pescado (rico en ácido eicosapentaenoico [EPA] y ácido docosahexaenoico [DHA]), aceite de microalgas (rico en DHA) y nucleótidos en el rendimiento reproductivo y del efecto subsiguiente en la calidad de los juveniles.”
A principios de abril del 2020, se transfirieron 180 parejas de lobina (M. salmoides, subespecie norteña) desde estanques de tierra a un sistema de recirculación acuícola (RAS, por sus siglas en inglés) en las instalaciones de Dunn´s Fish Farm en Brinkley, Arkansas.
El RAS cuenta con tres tanques (11.3 toneladas cada uno), unidad de biofiltro, filtro mecánico de tambor, intercambiador de calor, aireación y oxígeno puro que se adicionaba para garantizar que se mantuvieran niveles adecuados de oxígeno disuelto (Figura 1).

Sesenta parejas de reproductores se colocaron en cada tanque en condiciones de calidad de agua controladas (temperatura 17.9 ± 0.7°C, oxígeno disuelto 8.0 ± 0.5 mg/L, saturación de oxígeno disuelto 83.1 ± 4.5%) y fotoperíodo (10 h luz: 14 h oscuridad).
Después de aclimatar a los reproductores, se asignó aleatoriamente a los tanques uno de tres tratamientos dietéticos. Un grupo recibió una dieta comercial (48% proteína cruda, 18% lípidos crudos) como control (Dieta 1); las otras dos dietas se rociaron con aceite de pescado (0.5%) y aceite de microalgas (0.5%) (Dieta 2), y la Dieta 3 también se roció como la Dieta 2, pero también incluyó un nucleótido comercial (400 mg/kg).
La composición proximal y de ácidos grasos de las dietas se midió de acuerdo con los métodos 996.06 de la AOAC (2006). Al final del estudio de 92 días, los reproductores consumieron cantidades similares de dietas: Dieta 1, 105.3; Dieta 2, 102.5 y Dieta 3, 104.1 kg.
Posteriormente, los reproductores se transfirieron a tanques tipo pila de concreto para el desove donde la temperatura se incrementó de 17.8 a 20.3°C y el fotoperíodo se incrementó a 12 h de luz y 12 h de oscuridad. En los tanques de desove se colocaron nidos hechos con fibra de coco (45.7 cm × 45.7 cm) (Figura 2).

Cuando se presentaron los desoves, se retiraron los nidos y se colocaron en tanques de incubación (350 L), separados por tratamiento dietético (Figura 3).

Se tomaron muestras de huevos fertilizados y se realizaron mediciones del diámetro del huevo y del glóbulo de aceite con un microscopio (n = 50). Los huevos fertilizados se incubaron durante cinco días antes de que las larvas comenzaran a eclosionar.
Después de la eclosión, se llevó a cabo una prueba de inanición, se transfirieron 50 larvas a recipientes individuales (0.5 L) por triplicado de cada tratamiento dietético para evaluar la supervivencia a lo largo del tiempo cuando no se proporciona alimento (Figura 4). Se realizaron observaciones diarias de mortalidad.

Las larvas restantes se mantuvieron en tanques de incubación durante diez días adicionales antes de transferirlos a tanques más grandes de 1,000 L con densidad de 30 larvas/L (Figura 5).

A lo largo de este período, las larvas fueron alimentadas con rotíferos Brachionus plicatilis y Artemia salina (nauplios y metanauplios), y se destetaron gradualmente con alimento formulado en micropartícula (Otohime B1 y Otohime B2); luego solo se usó Otohime C1, antes de finalmente hacer la transición a alimento granulado.
El tamaño de los gránulos se incrementó de acuerdo con el crecimiento de los juveniles, de 0.8 a 1.2, 1.8, 2.5 hasta 3.5 mm. Después de ocho semanas, los juveniles se cosecharon y se registraron los parámetros de producción y crecimiento.
“La fecundidad y tasas de eclosión no se cuantificaron porque no pudimos rastrear de forma individual a las hembras. Sin embargo, el mayor número de larvas de reproductores alimentados con la Dieta 2 o 3 sugieren una mejor fecundidad y/o tasa de eclosión.”
Esta observación podría vincularse a huevos significativamente más grandes de los peces alimentados con la Dieta 3. De manera similar, el diámetro de los glóbulos de aceite fue significativamente mayor en los huevos producidos por los peces alimentados con las Dietas 2 o 3.
Para evaluar mejor la calidad de las larvas, se realizó la prueba de inanición. En el control, se produjo una mortalidad del 50% después de 7 días y una mortalidad del 100% después de 10 días. En cambio, se requirió más tiempo para que larvas murieran en los grupos de las Dietas 2 y 3.
Se produjo una mortalidad del 50% después de 9 días en los tratamientos de dietas suplementadas, mientras que la mortalidad total se produjo después de 11 y 15 días en la Dieta 3 y 2, respectivamente (Figura 6).
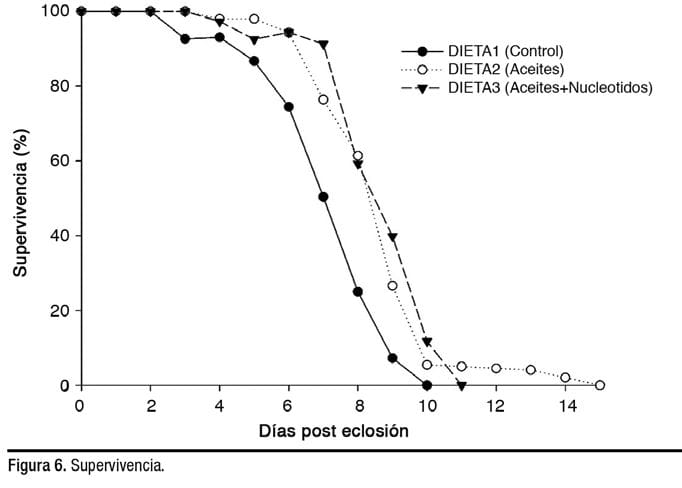
Tomados en conjunto, estos resultados indican que los reproductores hembra alimentados con las Dietas 2 o 3 depositaron más nutrientes en los huevos. La mayor disposición de nutrientes podría haber sido un factor decisivo en el mayor número larvas obtenidas de huevos producidos por los peces alimentados con la Dieta 2 (150,324) y la Dieta 3 (129,422), en comparación con la Dieta 1 (57,129).
Sin embargo, una vez que las larvas comenzaron la alimentación exógena, parece que la influencia de las dietas de los reproductores se minimizó en función del rendimiento del crecimiento (Figura 7).
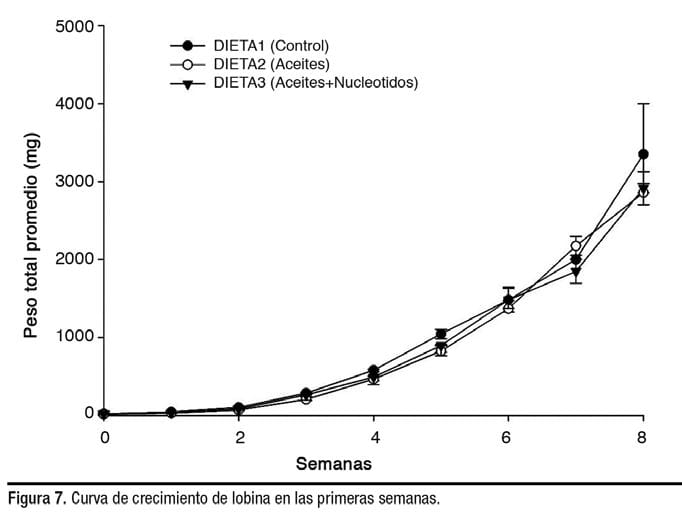
Las diferencias en el crecimiento de las larvas de la Dieta 1 durante las primeras semanas podrían deberse a las densidades más bajas de población y, por lo tanto, a una menor competencia por el alimento y el espacio, lo que permitió un peso individual más alto (0.026 mg) en comparación con las larvas de las Dietas 2 y 3 (0.012 mg cada uno).
Aunque la tasa de supervivencia de larvas en las Dietas 2 y 3 en la fase de destete fue menor (63.0 y 65.4%, respectivamente), la producción total de juveniles fue 175 y 156% mayor que la de los juveniles producidos con la Dieta 1.

Durante el ciclo de producción hubo varios casos de deformidades de la columna y malformación de la vejiga natatoria, en larvas (Figura 8) y juveniles (Figura 9). Esto probablemente fue un factor importante que contribuyó a la mortalidad, que incluyó tanto lordosis (curvatura ventral, forma de V) como cifosis (curvatura dorsal, forma de V invertida).

La condición de los reproductores y la calidad de los desoves son factores que afectan el desarrollo temprano de las células esqueléticas, el cartílago y/o la mineralización ósea en su progenie (Boglione et al., 2013a).
La nutrición de las larvas ha sido ampliamente reconocida como un parámetro clave que afecta la esqueletogénesis durante el desarrollo temprano de los peces, en particular, las deficiencias y/o desequilibrios en algunos nutrientes (lípidos, aminoácidos, vitaminas y minerales) (Boglione et al., 2013b).
“Por otra parte, el fallo y/o retraso en el inflado de la vejiga natatoria también puede provocar deformidades relacionadas con una hiperactividad de las larvas para mantener su posición en la columna de agua.”
Además de la nutrición y velocidad de la corriente del agua, los factores que afectan el inflado inicial de la vejiga natatoria pueden incluir la saturación gases, fotoperíodo, intensidad de la luz, temperatura y salinidad. Durante este estudio piloto, dos observaciones fueron claras:
1) las larvas malformadas tenían vejigas natatorias aparentemente bien infladas, y
2) estas patologías fueron menos comunes en juveniles de reproductores alimentados con las Dietas 2 y 3.
Estas observaciones preliminares requerirán un estudio más completo con repeticiones adicionales.
Sin embargo, se puede recomendar que, en los criaderos de lobina, se incremente el contenido de ácidos grasos poliinsaturados de cadena larga, especialmente DHA, mediante la suplementación con aceite de pescado y/o aceite de microalgas en una dieta comercial (48% de proteína bruta, 18% de lípidos brutos), como un enfoque sencillo para mejorar la producción de juveniles de lobina.
El FCA de la lobina y su mito en México
El cultivo de la lobina se encuentra poco desarrollado en México, a pesar de que existe alta demanda para consumo humano y de ser el pez más importante de pesca deportiva continental. La lobina es un pez carnívoro de alto nivel trófico (Brown et al., 2009), con reputación voraz, por su ímpetu al ingerir su alimento y por hacerlo en grandes cantidades y tamaños.
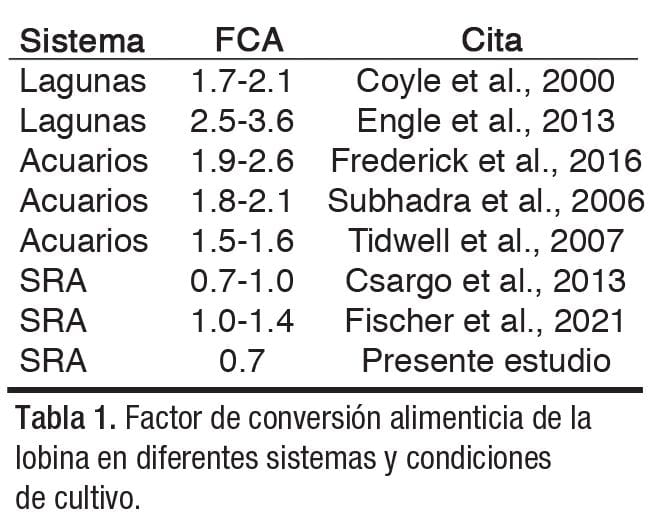
Quizá dicha reputación y muchos reportes no formales sobre su crianza, ha dado origen al mito de que en cautiverio presenta un factor de conversión alimenticia (FCA) exageradamente alto de hasta 10, lo que sugiere que se requieren 10 kilogramos de alimento para producir 1 kilogramo de lobina cultivada, poniendo en duda la sustentabilidad, rentabilidad y factibilidad de su cultivo.
Además, en México existen vacíos importantes en materia de investigación acuícola sobre este recurso. Aquí se presenta una revisión bibliográfica sobre valores de FCA de la lobina negra obtenidos desde diferentes sistemas y condiciones de cultivo (Tabla 1), así como la experiencia de los autores en el cultivo de la lobina boca grande, subespecie norteña.
Se llevó a cabo una evaluación de pre-engorda de juveniles por ocho semanas (marzo-abril, 2019), en el RAS que se describió en el apartado anterior. Se utilizaron 83,173 juveniles de 1.7 ± 0.2 g obtenidos del laboratorio (o del criadero)de la misma granja.
La densidad de siembra fue de 4.3 ± 0.8 kg·m-3 y/o 27,724 ± 967 peces por tanque. Al inicio, se alimentaron con pellets de 1.8 mm (Gemma Diamond; 57P:15L), en la tercer y quinta semana de evaluación se cambió a pellets de 2.5 y 3.5 mm, respectivamente (Biovita Fry; 50P:22L).
“La alimentación se realizó cada tres horas durante el fotoperiodo 15:9, a saciedad y registrándose la cantidad de alimento administrado. Semanalmente se registró peso y longitud totales de 50 peces por tanque para determinar factor de condición (k).”
También se determinó supervivencia, peso final, biomasa ganada, curva de crecimiento y FCA. Tres veces por día se registraron las variables fisicoquímicas del agua. La concentración de oxígeno fue de 7.5 ± 1.5 mg·L-1, temperatura 23.0 ± 1.0°C, pH 7.3 ± 0.1 y NH3 de 0.02 ± 0.02 mg·L-1.
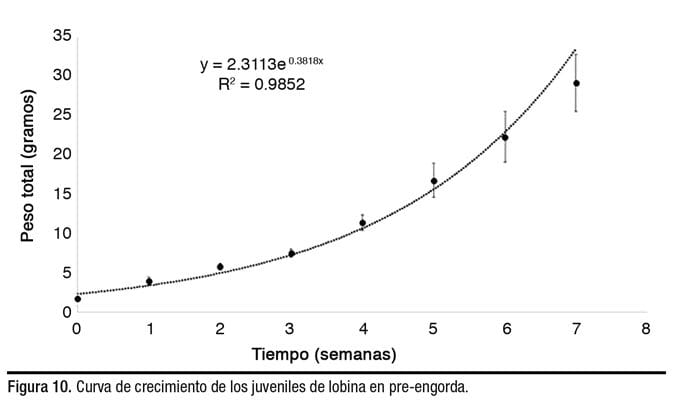
El valor de k a la octava semana fue de 1.6 ± 0.1. Se obtuvo una supervivencia de 97.7 ± 2.0%, peso final 28.1 ± 2.5 g, peso ganado de 713.7 ± 87.9 kg por tanque y FCA de 0.7 ± 0.03. Los valores de crecimiento semanal se ajustaron (R2 = 0.97) al modelo exponencial y= 1.5777e0.3818x (Figura 10).
Los resultados obtenidos muestran variables de producción aceptables para el cultivo de peces, el valor 0.7 de FCA registrado en la presente evaluación indica que la lobina tiene la capacidad de aprovechar eficientemente los alimentos administrados.
Además, la información bibliografía muestra que, en diferentes condiciones y sistemas de cultivo, la lobina en engorda presenta valores de 0.7 a 3 de FCA.
Por lo tanto, el supuesto alto FCA de la lobina es solo un mito, y en México se deben de dedicar mayores esfuerzos en materia de acuicultura para aprovechar al máximo este importante recurso.
Las referencias y fuentes consultadas por el autor en la elaboración de este artículo están disponibles bajo petición previa a nuestra redacción.
Ángel Humberto Rojo-Cebreros*
Mirian Celene León-López
Dunn’s Market Fish, Inc.
1078 Carson Brigde Rd.
Lonoke, AR 72086
*a.cebreros@dunnsmarketfish.com y arojocebreros@gmail.com




